
Why is nucleotide biosynthesis important?
Nucleotide biosynthesis is a highly regulated process necessary for cell growth and replication.
Where does nucleotide synthesis occur?
De novo purine nucleotide synthesis occurs actively in the cytosol of the liver where all of the necessary enzymes are present as a macro-molecular aggregate. The first step is a replacement of the pyrophosphate of PRPP by the amide group of glutamine.
Why is nucleotide metabolism important?
Nucleotide metabolism plays a central role in bacterial physiology, producing the nucleic acids necessary for DNA replication and RNA transcription.
What is biosynthesis of purine?
Purine biosynthesis requires ten enzymatic transformations to generate inosine monophosphate. PurF, PurD, PurL, PurM, PurC, and PurB are common to all pathways, while PurN or PurT, PurK/PurE-I or PurE-II, PurH or PurP, and PurJ or PurO catalyze the same steps in different organisms.
What are 3 types of nucleotide?
The three parts of a nucleotide are the nitrogenous base, pentose sugar, and phosphate group. The three parts of a nucleotide are the base, the sugar, and the phosphate. Nucleotides are the building blocks of DNA (2′-deoxyribonucleic acid) and RNA (ribonucleic acid).
What enzyme synthesizes nucleotides?
The enzyme that synthesizes DNA, DNA polymerase, can only add nucleotides to an already existing strand or primer of DNA or RNA that is base paired with the template. An enzyme, DNA polymerase, is required for the covalent joining of the incoming nucleotide to the primer.
What are the 4 functions of nucleotides?
Nucleotide FunctionNucleotides are the building block of DNA and RNA. ... Nucleotides act as coenzymes, which are required to catalyse many biochemical reactions by enzymes.Energy is stored in our body as ATP. ... NAD, NADP has an essential role to play in many redox reactions, they act as an electron carrier.More items...
What are examples of nucleotide?
A nucleotide is a monomer that serves as the building blocks for deoxyribonucleic acid (DNA) or ribonucleic acid (RNA). Examples of nucleotides are ribonucleotides or deoxyribonucleotides.
How does the body produce nucleotides?
After a meal, your pancreas secretes two types of enzymes, deoxyribonucleases, which break down DNA, and ribonucleases, which break down RNA. These enzymes cleave the DNA or RNA from your food into shorter chains of nucleotides, which your body then absorbs and transports to your cells for use.
What is biosynthesis of purine and pyrimidine?
Purine and pyrimidine nucleotides are major energy carriers, subunits of nucleic acids and precursors for the synthesis of nucleotide cofactors such as NAD and SAM. Despite the obvious importance of these molecules, we still have much to learn about how these nucleotides are synthesized and metabolized by plants.
How purine nucleotides are synthesized?
Purine synthesis is a ten-step process that requires ribose-5-phosphate from the PPP, glycine and formate from the serine/glycine synthesis pathway, glutamine, and TCA cycle-derived aspartate.
What purine means?
(PYOOR-een) One of two chemical compounds that cells use to make the building blocks of DNA and RNA. Examples of purines are adenine and guanine. Purines are also found in meat and meat products. They are broken down by the body to form uric acid, which is passed in the urine.
How are DNA nucleotides synthesized?
Synthesis of nucleotides requires a source of ribose 5-phosphate. This compound is produced from glucose 6-phosphate via the pentose phosphate pathway (also called the hexose monophosphate shunt).
How are nucleic acids synthesized?
Viral nucleic acid synthesis is catalyzed by both viral and host enzymes, the relative contribution of which is determined by the type of virus and the specific molecule. Viruses with RNA genomes, except for the retroviruses, synthesize mRNA and replicate their genomes using virus-encoded RNA-dependent RNA polymerases.
Why does the synthesis of nucleic acids occur in the direction of 5 → 3?
Why Does DNA Replication Go from 5' to 3'? DNA replication occurs in the 5' to 3' direction. DNA polymerase can only add nucleotides to the 3' OH group of the growing DNA strand, this is why DNA replication occurs only in the 5' to 3' direction.
How does the body synthesize amino acids?
All amino acids are synthesized from intermediates in glycolysis, the citric acid cycle, or the pentose phosphate pathway. Nitrogen is provided by glutamate and glutamine. Amino acid synthesis depends on the formation of the appropriate alpha-keto acid, which is then transaminated to form an amino acid.
What are the glycoconjugates of trypanosomatids?
The biosynthesis of trypanosomatid glycoconjugates is closely linked to central carbon metabolism and the synthesis of sugar nucleotides and dolichol (Dol)-linked donors that are utilized by glycosyltransferases in both the secretory pathway and cytoplasm. Consistent with the known glycan structures synthesized by different trypanosomatids, all of these parasites have the capacity to synthesize the sugar nucleotides UDP-Glc, UDP-Gal, UDP-GlcNAc, GDP-Man, and GDP-Fuc88 ( Fig. 4 ). In addition, T. cruzi synthesize UDP-Gal f, UDP-Rha, and UDP-Xyl, while Leishmania spp. synthesize UDP-Gal f and GDP-D-Ara p88 ( Fig. 4 ). While many of the enzymes involved in sugar nucleotide biosynthesis are conserved across all trypanosomatids, significant genera-specific differences occur in the extent to which these parasites can directly scavenge extracellular sugars and convert them to sugar nucleotides, versus synthesis by interconversion of other sugar nucleotides ( Fig. 4 ). Distinctive features of sugar nucleotide biosynthesis in each of the major trypanosomatid groups are summarized below.
What enzyme is responsible for the conversion of dihydroorotate to carbamoyl aspartate?
Dihydroorotase (DHO) catalyzes the reversible hydrolytic conversion of dihydroorotate and carbamoyl aspartate (Figure 6 (e)), an important step in pyrimidine nucleotide biosynthesis. The crystal structure of DHO from E. coli 169 has been solved and illuminates an overall fold described as a (β/α) 8 -barrel indicating that this enzyme is a member of the amidohydrolase superfamily. The active site of DHO contains a binuclear metal center that is occupied by two Zn 2 + ions separated by 3.6 Ǻ. The zinc ion bound to the Mα site is coordinated by the side chains of His16, His18, Lys102, Asp250 and a solvent molecule in a trigonal bipyramidal geometry. The zinc ion bound at the M β site is coordinated by Lys102, His139, His177 and a water molecule in tetrahedral geometry. The bridging ligands for the two zinc ions are Lys102 (carbamoylated) and the solvent molecule. Other members of the amidohydrolase superfamily share a similar binuclear metal ligation and proposed reaction mechanisms, including phosphotriesterase (PTE), iso-aspartyl dipeptidase (IAD) and possibly urease (URE). The binuclear metal centers for these enzymes are occupied by either Zn2 + (PTE, IAD) or Ni 2 + (URE). 30
Where is the de novo nucleotide synthesis site?
Within the body the major site of de novo nucleotide synthesis, for the replenishment and maintenance of intracellular pools, is the liver . Following their synthesis in the liver the nucleotides are dephosphorylated and in part phosphorolytically cleaved into nucleobases and ribose-1-phosphate for transport to the blood and then subsequent uptake by cells of the other organs.
What happens to nucleosides during catabolism?
During the catabolism of nucleic acids, nucleoside mono- and diphosphates are released. The nucleosides do not accumulate to any significant degree, owing to the action of nucleoside kinases. These include both nucleoside monophosphate (NMPK) kinases and nucleoside diphosphate (NDPK) kinases.
What is the pathway of purine and pyrimidine?
Both the salvage and de novo synthesis pathways of purine and pyrimidine biosynthesis lead to production of nucleoside-5′-phosphates through the utilization of an activated sugar intermediate and a class of enzymes called phosphoribosyltransferases. The activated sugar used is 5-phosphoribosyl-1-pyrophosphate, PRPP. PRPP is generated by the action of PRPP synthetase (also called ribose-phosphate pyrophosphokinase 1) and requires energy in the form of ATP.
What is the catabolism of purine nucleotides?
The catabolism of purine nucleotides involves deamination reaction, phosphate removal from the nucleoside monophosphates, phosphorylytic removal of the ribose yielding ribose-1-phosphate, and finally oxidation of the nucleobases to uric acid.
What is the catabolic reaction of pyrimidine nucleotides?
Catabolism of the pyrimidine nucleotides leads ultimately to β-alanine (when CMP and UMP are degraded) or β-aminoisobutyrate (when dTMP is degraded) and NH 3 and CO 2. The β-alanine and β-aminoisobutyrate serve as -NH 2 donors in transamination of α-ketoglutarate to glutamate. A subsequent reaction converts the products to malonyl-CoA (which can be diverted to fatty acid synthesis) or methylmalonyl-CoA (which is converted to succinyl-CoA and can be shunted to the TCA cycle).
How are bases hydrolyzed?
The bases are hydrolyzed from nucleosides by the action of phosphorylases that yield ribose-1-phosphate and free nucleobases. If the nucleosides and/or bases are not re-utilized the purine bases are further degraded to uric acid and the pyrimidines to β-aminoiosobutyrate, NH 3 and CO 2.
What enzymes are responsible for the hydrolysis of nucleic acids?
Extracellular hydrolysis of ingested nucleic acids occurs through the concerted actions of endonucleases, phosphodiesterases, and nucleoside phosphorylases. Endonucleases degrade DNA and RNA at internal sites leading to the production of oligonucleotides. Oligonucleotides are further digested by phosphodiesterases that act from the ends inward yielding free nucleosides. The bases are hydrolyzed from nucleosides by the action of phosphorylases that yield ribose-1-phosphate and free nucleobases. If the nucleosides and/or bases are not re-utilized the purine bases are further degraded to uric acid and the pyrimidines to β-aminoiosobutyrate, NH 3 and CO 2.
What are nucleotides in metabolism?
Nucleotides also act in metabolic regulation, as in the response of key enzymes of intermediary metabolism to the relative concentrations of AMP, ADP, and ATP (PFK is a prime example here; see also Chapter 19 ).
How many enzymes are involved in pyrimidine biosynthesis?
In bacteria, the six enzymes of de novo pyrimidine biosynthesis exist as distinct proteins, each independently catalyzing its specific step in the overall pathway. In contrast, in mammals, the six enzymatic activities are distributed among only three proteins, two of which are multifunctional polypeptides: single polypeptide chains having two or more enzymic centers. The first three steps of pyrimidine synthesis, CPS-II, aspartate transcarbamoylase, and dihydroorotase, are all localized on a single 210-kD cytosolic polypeptide. This multifunctional enzyme is the product of a solitary gene, yet it is equipped with the active sites for all three enzymatic activities. Step 4 ( Figure 27.17) is catalyzed by DHO dehydrogenase, a separate enzyme associated with the outer surface of the inner mitochondrial membrane, but the enzymatic activities mediating Steps 5 and 6, namely, orotate phosphoribosyltransferase and OMP decarboxylase in mammals, are also found on a single cytosolic polypeptide known as UMP synthase.#N#The purine biosynthetic pathway of avian liver also provides examples of metabolic channeling. Recall that Steps 3, 4, and 6 of de novo purine synthesis are catalyzed by three enzymatic activities localized on a single multifunctional polypeptide, and Steps 7 and 8 and Steps 10 and 11 by respective bifunctional polypeptides (see Figure 27.3 ).
How does thymine synthesis occur?
The synthesis of thymine nucleotides proceeds from other pyrimidine deoxyribonucleotides. Cells have no requirement for free thymine ribonucleotides and do not synthesize them. Small amounts of thymine ribonucleotides do occur in tRNA (an RNA species harboring a number of unusual nucleotides), but these Ts arise via methylation of U residues already incorporated into the tRNA. Both dUDP and dCDP can lead to formation of dUMP, the immediate precursor for dTMP synthesis (Figure 27.27). Interestingly, formation of dUMP from dUDP passes through dUTP, which is then cleaved by dUTPase, a pyrophosphatase that removes PP i from dUTP. The action of dUTPase prevents dUTP from serving as a substrate in DNA synthesis. An alternative route to dUMP formation starts with dCDP, which is dephosphorylated to dCMP and then deaminated by dCMP deaminase (Figure 27.28), leaving dUMP. dCMP deaminase provides a second point for allosteric regulation of dNTP synthesis; it is allosterically activated by dCTP and feedback-inhibited by dTTP. Of the four dNTPs, only dCTP does not interact with either of the regulatory sites on ribonucleotide reductase (Figure 27.26). Instead, it acts upon dCMP deaminase.
What is the de novo pathway for purine synthesis?
Figure 27.3 · The de novo pathway for purine synthesis. The first purine product of this pathway, IMP (inosinic acid or inosine monophosphate), serves as a precursor to AMP and GMP. Step 1: PRPP synthesis from ribose-5-phosphate and ATP by ribose-5-phosphate pyrophosphokinase. Step 2: 5-Phosphoribosyl-b-1-amine synthesis from a-PRPP, glutamine, and H 2 O by glutamine phosphoribosyl pyrophosphate amidotransferase. Step 3: Glycinamide ribonucleotide (GAR) synthesis from glycine, ATP, and 5-phosphoribosyl-b-amine by glycinamide ribonucleotide synthetase. Step 4: Formylglycinamide ribonucleotide synthesis from N10 -formyl-THF and GAR by GAR transformylase. Step 5: Formylglycinamidine ribonucleotide (FGAM) synthesis from FGAR, ATP, glutamine, and H 2 O by FGAM synthetase (FGAR amidotransferase). The other products are ADP, P i, and glutamate. Step 6: 5-Aminoimidazole ribonucleotide (AIR) synthesis is achieved via the ATP-dependent closure of the imidazole ring, as catalyzed by FGAM cyclase (AIR synthetase). (Note that the ring closure changes the numbering system.) Step 7: Carboxyaminoimidazole ribonucleotide (CAIR) synthesis from CO 2, ATP, and AIR by AIR carboxylase. Step 8: N-succinylo-5-aminoimidazole-4-carboxamide ribonucleotide (SAICAR) synthesis from aspartate, CAIR, and ATP by SAICAR synthetase. Step 9: 5-Aminoimidazole carboxamide ribonucleotide (AICAR) formation by the nonhydrolytic removal of fumarate from SAICAR. The enzyme is adenylosuccinase. Step 10: 5-Formylaminoimidazole carboxamide ribonucleotide (FAICAR) formation from AICAR and N 10 -formyl-THF by AICAR transformylase. Step 11: Dehydration/ring closure yields the authentic purine ribonucleotide IMP. The enzyme is IMP synthase.
What is the step of ribose 5 phosphate?
In Step 1 , ribose-5-phosphate is activated via the direct transfer of a pyrophosphoryl group from ATP to C-1 of the ribose, yielding 5-phosphoribosyl-a-pyrophosphate (PRPP) (Figure 27.3). The enzyme is ribose-5-phosphate pyrophosphokinase. PRPP is the limiting substance in purine biosynthesis.
Which site binds ATP?
The second effector site, the substrate specificity site, can bind either ATP, dTTP, dGTP, or dATP, and the substrate specificity of the enzyme is determined by which of these nucleotides occupies this site. If ATP is in the substrate specificity site, ribonucleotide reductase preferentially binds pyrimidine nucleotides (UDP or CDP) at its active site and reduces them to dUDP and dCDP. With dTTP in the specificity-determining site, GDP is the preferred substrate. When dGTP binds to this specificity site, ADP becomes the favored substrate for reduction. The rationale for these varying affinities is as follows (Figure 27.26): High [ATP] is consistent with cell growth and division and, consequently, the need for DNA synthesis. Thus, ATP binds in the activity-determining site of ribonucleotide reductase, turning it on and promoting production of dNTPs for DNA synthesis. Under these conditions, ATP is also likely to occupy the substrate specificity site, so that UDP and CDP are reduced to dUDP and dCDP. As we shall soon see, both of these pyrimidine deoxynucleoside diphosphates are precursors to dTTP. Thus, elevation of dUDP and dCDP levels leads to an increase in [dTTP]. High dTTP levels increase the likelihood that it will occupy the substrate specificity site, in which case GDP becomes the preferred substrate, and dGTP levels rise. Upon dGTP association with the substrate specificity site, ADP is the favored substrate, leading to ADP reduction and the eventual accumulation of dATP. Binding of dATP to the overall activity site then shuts the enzyme down. In summary, the relative affinities of the three classes of nucleotide binding sites in ribonucleotide reductase for the various substrates, activators, and inhibitors are such that the formation of dNDPs proceeds in an orderly and balanced fashion. As these dNDPs are formed in amounts consistent with cellular needs, their phosphorylation by nucleoside diphosphate kinases produces dNTPs, the actual substrates of DNA synthesis.
How is DTMP synthesized?
Synthesis of dTMP from dUMP is catalyzed by thymidylate synthase (Figure 27.29). This enzyme methylates dUMP at the 5-position to create dTMP; the methyl donor is the one-carbon folic acid derivative N5, N10 -methylene-THF. The reaction is actually a reductive methylation in which the one-carbon unit is transferred at the methylene level of reduction and then reduced to the methyl level. The THF cofactor is oxidized at the expense of methylene reduction to yield dihydrofolate, or DHF. Dihydrofolate reductase then reduces DHF back to THF for service again as a one-carbon vehicle (see Figure 18.35 ). Thymidylate synthase sits at a junction connecting dNTP synthesis with folate metabolism. It has become a preferred target for inhibitors designed to disrupt DNA synthesis. An indirect approach is to employ folic acid precursors or analogs as antimetabolites of dTMP synthesis (Figure 27.30). Purine synthesis is affected as well because it is also dependent on THF ( Figure 27.3 ).
What is the effect of inhibiting the synthesis of dexoyribonucleotides?
Inhibition of the synthesis of dexoyribonucleotides or thymidylate will selectively inhibit fast growing cells.
Which radical recaptures the hydrogen that was abstracted by the first cysteine residue?
5. The C–3' radical recaptures the hydrogen that was abstracted by the first cysteine residue.
What is the reducing agent of ribose sugar?
The 2'-OH on the ribose sugar is reduced to a hydrogen. NADPH + H+is the reducing agent. The enzyme is called ribonucleotide reductase
How many phosphate groups are in ribose sugar?
Ribose or deoxyribose sugar One to three phosphate groups purine or pyrimidine hetercyclic nitrogen base.
What is the reaction driven by?
Reaction is driven by the hydorlysis of pyrophosphate
Which radical promotes the release of a hydorxide ion on carbon-2?
3. The carbon radical at C-3' promotes the release of a hy dorxide ion on carbon-2.
What is converted to UTP before going on to produce CTP?
Nucleoside monophosphate kinases UMP is converted to UTP before going on to produce CTP
What are the roles of nucleotides in biosynthesis?
Biosynthesis of nucleotides. Nucleotides play a variety of important roles in all cells. They are the precursors of DNA and RNA. They are essential carriers of chemical energy—a role primarily of ATP and to some extent GTP. They are components of the cofactors NAD, FAD, S-adenosylmethionine, and coenzyme A, as well as of activated biosynthetic ...
How does pyrimidine nucleotide synthesis occur in bacteria?
Regulation of the rate of pyrimidine nucleotide synthesis in bacteria occurs in large part through aspartate transcarbamoylase (ATCase), which catalyzes the first reaction in the sequence and is inhibited by CTP, the end product of the sequence (Fig. 16–4a). The bacterial ATCase molecule consists of six catalytic subunits and six regulatory subunits. The catalytic subunits bind the substrate molecules, and the allosteric subunits bind the allosteric inhibitor, CTP. The entire ATCase molecule, as well as its subunits, exists in two conformations, active and inactive. When CTP is not bound to the regulatory subunits, the enzyme is maximally active. As CTP accumulates and binds to the regulatory subunits, they undergo a change in conformation. This change is transmitted to the catalytic subunits, which then also shift to an inactive conformation. ATP prevents the changes induced by CTP.
How is ribonucleotide reductase regulated?
coli ribonucleotide reductase is unusual in that not only its activity but its substrate specificity is regulated by the binding of effector molecules. Each R1 subunit has two types of regulatory site. One type affects overall enzyme activity and binds either ATP, which activates the enzyme, or dATP, which inactivates it. The second type alters substrate specificity in response to the effector molecule— ATP, dATP, dTTP, or dGTP—that is bound there (Fig. 16–8). When ATP or dATP is bound, reduction of UDP and CDP is favored. When dTTP or dGTP is bound, reduction of GDP or ADP, respectively, is stimulated. The scheme is designed to provide a balanced pool of precursors for DNA synthesis. ATP is also a general activator for biosynthesis and ribonucleotide reduction. The presence of dATP in small amounts increases the reduction of pyrimidine nucleotides. An oversupply of the pyrimidine dNTPs is signaled by high levels of dTTP, which shifts the specificity to favor reduction of GDP. High levels of dGTP, in turn, shift the specificity to ADP reduction, and high levels of dATP shut the enzyme down. These effectors are thought to induce several distinct enzyme conformations with altered specificities.
How is pyrimidine synthesized?
The pyrimidine is constructed from carbamoyl phosphate and aspartate. The ribose 5-phosphate is then added to the completed pyrimidine ring by orotate phosphoribosyltransferase. The first step in this pathway (not shown here; is the synthesis of carbamoyl phosphate from CO2 and NH4+, catalyzed in eukaryotes by carbamoyl phosphate synthetase II. (b) Channeling of intermediates in bacterial carbamoyl phosphate synthetase. (Derived from PDB ID 1M6V.) The large and small subunits are shown in gray and blue, respectively; the channel between active sites (almost 100 Å long) is shown as a yellow mesh. A glutamine molecule (green) binds to the small subunit, donating its amido nitrogen as NH4+ in a glutamine amidotransferase–type reaction. The NH4+ enters the channel, which takes it to a second active site, where it combines with bicarbonate in a reaction requiring ATP (bound ADP in blue). The carbamate then reenters the channel to reach the third active site, where it is phosphorylated to carbamoyl phosphate (bound ADP in red).
Where is carbamoyl phosphate synthesized?
However, in animals the carbamoyl phosphate required in urea synthesis is made in mitochondria by carbamoyl phosphate synthetase I, whereas the carbamoyl phosphate required in pyrimidine biosynthesis is made in the cytosol by a different form of the enzyme, carbamoyl phosphate synthetase II.
How are deoxyribonucleotides derived from DNA?
Deoxyribonucleotides, the building blocks of DNA, are derived from the corresponding ribonucleotides by direct reduction at the 2'-carbon atom of the D-ribose to form the 2'-de oxy derivative. For example, adenosine diphosphate (ADP) is reduced to 2'-deoxyadenosine-diphosphate (dADP), and GDP is reduced to dGDP. This reaction is somewhat unusual in that the reduction occurs at a nonactivated carbon; no closely analogous chemical reactions are known. The reaction is catalyzed by ribonucleotide reductase, best characterized in E. coli, in which its substrates are ribonucleoside diphosphates.#N#The reduction of the D-ribose portion of a ribonucleoside diphosphate to 2'-deoxy-D-ribose requires a pair of hydrogen atoms, which are ultimately donated by NADPH via an intermediate hydrogen-carrying protein, thioredoxin. This ubiquitous protein serves a similar redox function in photosynthesis and other processes. Thioredoxin has pairs of -SH groups that carry hydrogen atoms from NADPH to the ribonucleoside diphosphate. Its oxidized form (S-S) is reduced by NADPH in a reaction catalyzed by thioredoxin reductase (Fig. 16–5), and reduced thioredoxin is then used by ribonucleotide reductase to reduce the nucleoside diphosphates (NDPs) to deoxyribonucleoside diphosphates (dNDPs).#N#A second source of reducing equivalents for ribonucleotide reductase is glutathione (GSH). Glutathione serves as the reductant for a protein closely related to thioredoxin, glutaredoxin, which then transfers the reducing power to ribonucleotide reductase (Fig. 16–5). Ribonucleotide reductase is notable in that its reaction mechanism provides the best-characterized example of the involvement of free radicals in biochemical transformations, once thought to be rare in biological systems. The enzyme in E. coli and most eukaryotes is a dimer, with subunits designated R1 and R2 (Fig. 16–6). The R1 subunit contains two kinds of regulatory sites, as described below. The two active sites of the enzyme are formed at the interface between the R1 and R2 subunits. At each active site, R1 contributes two sulfhydryl groups required for activity and R2 contributes a stable tyrosyl radical. The R2 subunit also has a binuclear iron (Fe3+) cofactor that helps generate and stabilize the tyrosyl radicals. The tyrosyl radical is too far from the active site to interact directly with the site, but it generates another radical at the active site that functions in catalysis.
What are the steps of multifunctional enzymes?
In humans, a multifunctional enzyme combines the activities of AIR carboxylase and SAICAR synthetase (steps 6a and 8). In bacteria, these activities are found on separate proteins, but a large noncovalent complex may exist in these cells.
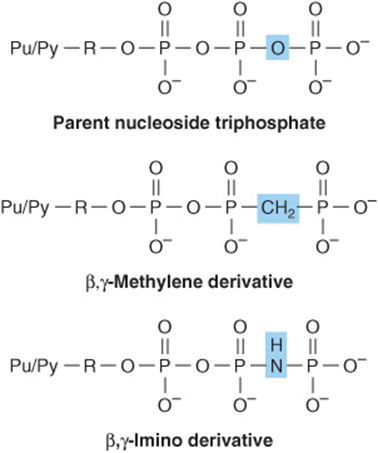
Introduction to Nucleic Acids
Activation of Ribose-5-Phosphate
- Both the salvage and de novo synthesis pathways of purine and pyrimidine biosynthesis lead to production of nucleoside-5′-phosphates through the utilization of an activated sugar intermediate and a class of enzymes called phosphoribosyltransferases. The activated sugar used is 5-phosphoribosyl-1-pyrophosphate, PRPP. PRPP is generated by the action of PRPP synthetase (…
Purine Nucleotide Biosynthesis
- Synthesis of Inosine Monophosphate: IMP
The major site of purine synthesis is in the liver. Synthesis of the purine nucleotides begins with PRPP and leads to the first fully formed nucleotide, inosine 5′-monophosphate (IMP). This pathway is diagrammed below. The purine base without the attached ribose moiety is hypoxanth… - Synthesis of AMP and GMP from IMP
IMP represents a branch point for purine biosynthesis, because it can be converted into either AMP or GMP through two distinct reaction pathways. The pathway leading to AMP requires energy in the form of GTP; that leading to GMP requires energy in the form of ATP. The utilizatio…
Regulation of Purine Nucleotide Synthesis
- The essential rate limiting steps in purine biosynthesis occur at the first two steps of the pathway. The synthesis of PRPP by PRPP synthetase is feed-back inhibited by purine-5′-nucleotides (predominantly AMP and GMP). Combinatorial effects of those two nucleotides are greatest, e.g., inhibition is maximal when the correct concentration of both adenine and guanine nucleotides is …
Catabolism of Purine Nucleotides
- Catabolism of the purine nucleotides (both ribonucleotides and deoxyribonucleotides) leads ultimately to the production of uric acid which is insoluble and is excreted in the urine. Uric acid excretion and reabsorption occurs within the proximal tubulesof the kidney. Elevation in uric acid levels can result in precipitation of urate crystals with monosodium urate crystals being the most …
Salvage of Purine Nucleotides
- The re-synthesis of nucleotides from the purine bases and purine nucleosides takes place in a series of steps known as the salvage pathways. The free purine bases, adenine, guanine, and hypoxanthine, can be reconverted to their corresponding nucleotides by phosphoribosylation where PRPP, like in the de novosynthesis pathway, serves as the activated form of ribose-5′-ph…
Clinical Significances of Purine Metabolism
- Clinical problems associated with nucleotide metabolism in humans are predominantly the result of abnormal catabolism of the purines. The clinical consequences of abnormal purine metabolism range from mild to severe and even fatal disorders. Clinical manifestations of abnormal purine catabolism arise from the insolubility of the degradation byproduct, uric acid. Gout is a condition …
Table of Disorders of Purine Metabolism
- ahypoxanthine-guanine phosphoribosyltransferase
- badenosine deaminase
- cpurine nucleotide phosphorylase
- dadenosine phosphoribosyltransferase
Pyrimidine Nucleotide Biosynthesis
- Synthesis of the pyrimidines is less complex than that of the purines, since the base is much simpler. The first completed base is derived from one mole of glutamine, one mole of ATP and one mole of CO2(which form carbamoyl phosphate) and one mole of aspartate. An additional mole of glutamine and ATP are required in the conversion of UTP to CTP. The pathway of pyrim…
Synthesis of The Thymine Nucleotides
- The de novo pathway to dTTP synthesis first requires the use of dUMP from the metabolism of either UDP or CDP. The dUMP is converted to dTMP (5-methyl-dUMP) by the action of thymidylate synthetase. The methyl group (recall that thymine is 5-methyl uracil) is donated by N5,N10-methylene THF. The unique property of the action of thymidylate synthetase is that i…