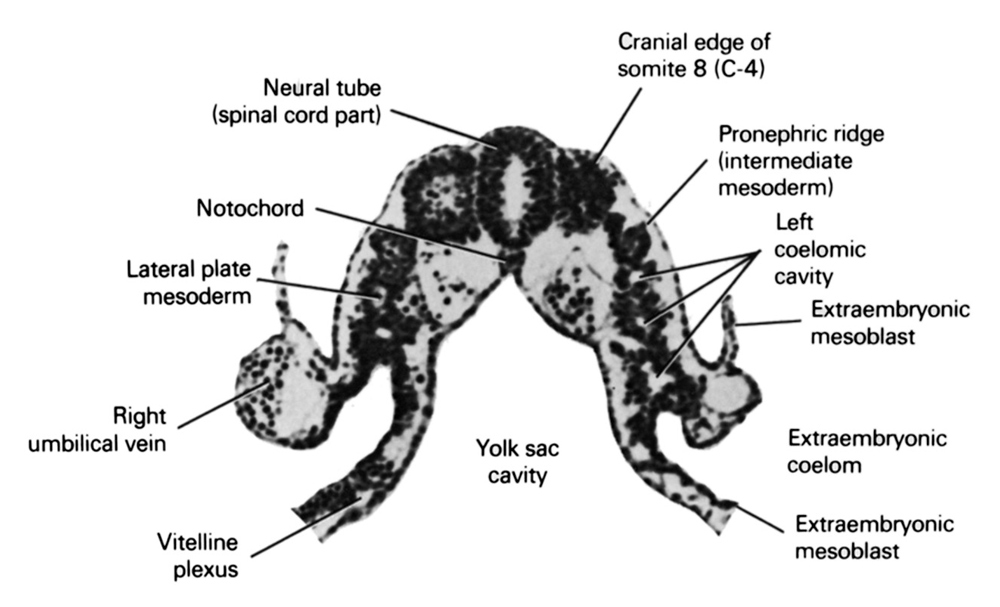
Out of the somites arise the sclerotome, forerunner of the bodies and neural arches of the vertebrae; the dermatome, precursor of the connective tissue of the skin; and the myotome, or primitive muscle, from which the major muscles of vertebrates are derived.
What are somites in vertebrates?
In vertebrates, somites subdivide into the sclerotomes, myotomes, syndetomes and dermatomes that give rise to the vertebrae of the vertebral column, rib cage, part of the occipital bone, skeletal muscle, cartilage, tendons, and skin (of the back). [2] The word somite is sometimes also used in place of the word metamere.
Which layer do somites give rise to?
- Somites give rise to cells that form vertebrae, ribs, most skeletal muscle and dermis of dorsal skin. Somites are derived from which layer? Mesoderm Invaginating epiblast cells that lie between developing ectoderm and endoderm will become mesoderm. - These three germ layers are the primordia for all tissues & organs in the adult.
What are the components of somitogenesis?
The important components of somitogenesis (somite formation) are periodicity, epithelialization, specification, and differentiation. The first somites appear in the anterior portion of the trunk, and new somites “bud off” from the rostral end of the paraxial mesoderm at regular intervals (Figures 14.2C,Dand 14.3).
What is the dorsal epithelial structure of the somite?
The dorsal epithelial structure of the somite is maintained in the dermomyotome which eventually gives rise to the epaxial muscles of the vertebrae and back (medial myotome), the hypaxial muscles of the body wall and limbs (lateral myotome), and the dermis of the skin of the trunk (dermatome). Figure 1.
How do somites form?
The important components of somitogenesis (somite formation) are periodicity, epithelialization, specification, and differentiation. The first somites appear in the anterior portion of the trunk, and new somites “bud off” from the rostral end of the paraxial mesoderm at regular intervals (Figures 14.2C,Dand 14.3). Somite formation begins as paraxial mesoderm cells become organized into whorls of cells called somitomeres. The somitomeres become compacted and bound together by an epithelium, and eventually separate from the presomitic paraxial mesoderm to form individual somites. Because individual embryos can develop at slightly different rates (as when chick embryos are incubated at slightly different temperatures), the number of somites present is usually the best indicator of how far development has proceeded. The total number of somites formed is characteristic of a species (50 in chicks, 65 in mice, and about 500 in some snakes).
What is the expression pattern of Somite X?
Somite X is marked. The expression of the hairygene (purple) is seen in the caudal half of this somite, (more...) Thus, the expression pattern of the hairygene correlates with the positioning of the place where a somite will separate from the unsegmented mesoderm.
What is the term for the segmental plate in birds?
These bands of paraxial mesoderm are referred to as the segmental plate (in birds) and the unsegmented mesoderm (in mammals). As the primitive streak regresses and the neural folds begin to gather at the center of the embryo, the paraxial mesoderm separates into blocks of cells called somites.
How are myotomes induced?
In similar ways, the myotome is induced by at least two distinct signals. Studies involving transplantation and knockout mice indicate that the epaxial muscle cells coming from the medial portion of the somite are induced by factors from the neural tube, probably Wnt1 and Wnt3a from the dorsal region and low levels of Sonic hedgehog from the ventral region (Münsterberg et al. 1995; Stern et al. 1995; Ikeya and Takada 1998). The hypaxial muscles coming from the lateral edge of the somite are probably induced by a combination of Wnt proteins from the epidermis and bone morphogenetic protein 4 (BMP4) from the lateral plate mesoderm (Cossu et al. 1996a; Pourquié et al. 1996; Dietrich et al. 1998). These factors cause the myotome cells to express particular transcription factors that activate the muscle-specific genes.
What is the position of the segmental plate mesoderm?
The segmental plate mesoderm is determined as to its position along the anterior-posterior axis before somitogenesis. When segmental plate mesoderm that would ordinarily form thoracic somites is transplanted into a region in a younger embryo (caudal to (more...) Differentiation within the somite.
Which layer of the dermamyotome is the central region of the dermamyot?
The central region of the dorsal layer of the dermamyotome is called the dermatome, and it generates the mesenchymal connective tissue of the back skin: the dermis. (The dermis of other areas of the body forms from other mesenchymal cells, not from the somites.)
Where does the head musculature come from?
14.2 Cranial paraxial mesoderm. Most of the head musculature does not come from somites. Rather, it comes from the cranial paraxial mesoderm. These cells originate adjacent to the sides of the brain, and they migrate to their respective destinations. http://www.devbio.com/chap14/link1402.shtml
What is a somite?
Somites are transient structures that do not exist in the adult. They disappear as organogenesis proceeds.
Which part of somite is the direct expression of Myf5 and MyoD?
from lateral mesoderm direct expression of Myf5 and MyoD in dorsolateral part of somite initiating limb and body wall muscles.
When does paraxial mesoderm begin to organize in segments?
paraxial mesoderm begins to organize in segments by Wk 3
How many primary sources does the skeleton develop from?
The skeleton develops from three primary sources?
Which epiblast cells lie between the ectoderm and endoderm?
Invaginating epiblast cells that lie between developing ectoderm and endoderm will become mesoderm.
When does mesoderm thicken?
The initial sheet of mesoderm thickens near midline by Day 17
What is the segmental heritage of vertebrates?
The segmental heritage of all vertebrates is evident in the character of the vertebral column. And yet, the extent to which direct translation of pattern from the somitic mesoderm and de novo cell and tissue interactions pattern the vertebral column remains a fundamental, unresolved issue. The elements of vertebral column pattern under debate include both segmental pattern and anteroposterior regional specificity. Understanding how vertebral segmentation and anteroposterior positional identity are patterned requires understanding vertebral column cellular and developmental biology. In this study, we characterized alignment of somites and vertebrae, distribution of individual sclerotome progeny along the anteroposterior axis and development of the axial skeleton in zebrafish. Our clonal analysis of zebrafish sclerotome shows that anterior and posterior somite domains are not lineage-restricted compartments with respect to distribution along the anteroposterior axis but support a ‘leaky’ resegmentation in development from somite to vertebral column. Alignment of somites with vertebrae suggests that the first two somites do not contribute to the vertebral column. Characterization of vertebral column development allowed examination of the relationship between vertebral formula and expression patterns of zebrafish Hox genes. Our results support co-localization of the anterior expression boundaries of zebrafish hoxc6 homologs with a cervical/thoracic transition and also suggest Hox-independent patterning of regionally specific posterior vertebrae.
What is the shift in register observed between somites and vertebral column?
The shift in register observed between somites and vertebral column is posited to occur when the posterior sclerotome of one somite pair recombines with the anterior sclerotome of the next posterior somite pair. The compartmental version of resegmentation holds that the anterior and posterior domains are lineage-restricted compartments. This conception of resegmentation has a direct correlate in the anterior and posterior compartments that subdivide ectodermal parasegments in D. melanogaster. But does this model describe how the vertebral column is derived from somites in vertebrate embryos?
What is the axial skeleton of a zebrafish?
Histological staining revealed that the zebrafish axial skeleton developed in a stereotyped sequence. We characterized primary elements of the axial skeleton, including the centrum, neural and hemal arches, and ribs. Because the rate at which zebrafish larvae developed was quite variable, the age at which various stages in the sequence of vertebral development were reached also varied considerably. Thus, ages given below represent the earliest age at which each stage in vertebral development was observed. The anterior end of the zebrafish notochord stains with Alizarin Red, ossifies and is surrounded by the developing basioccipital bone of the cranium ( Cubbage and Mabee, 1996 ). We found that anterior notochord was marked by matrix stains in concert with development of other elements of the cranium (data not shown). These events initiated approximately 3 days prior to the onset of stainable matrix in the post-cranial axial skeleton ( Cubbage and Mabee, 1996) (data not shown). Therefore, given both developmental timing and position within the embryo, we classify the matrix surrounding the anterior-most portion of the notochord as a head element as do Cubbage and Mabee ( Cubbage and Mabee, 1996) and Kimmel et al. ( Kimmel et al., 2001 ), but in contrast to a recent report by Du and colleagues ( Du et al., 2001) in which this region is labeled as the first element of the vertebral column. We found that the three most anterior vertebral centra were the first to develop cartilaginous matrix. Stained foci were evident dorsal to the notochord at 6 d with Alizarin Red (data not shown) and at 7 d with Alcian Blue ( Fig. 2 A). Later development of the two anterior-most centra lagged behind development of the remaining trunk and tail centra, a phenomenon also noted by Du and colleagues ( Du et al., 2001 ). Additional foci arose ventrally and eventually rings of smoothly stained matrix surrounded the notochord ( Fig. 2 B) ( Fleming et al., 2001 ). Development of the centra proceeded in an anterior to posterior sequence ( Du et al., 2001 ), apparent with both Alcian Blue and Alizarin Red, and was complete along the length of the fish at 9 d. Neural and hemal arches on individual vertebrae arose synchronously in an anterior to posterior sequence first visible at 8 d by α-coll2a1 expression (data not shown) and at 11 d with Alcian Blue staining. In contrast to the smooth appearance of the stain surrounding the notochord, the chondrocytes of the developing neural arches were readily apparent ( Fig. 2 C). The general anterior to posterior sequence of development was retained for the centra, neural and hemal arches, and rib development. At 21 d, the axial skeleton was essentially complete with evidence of all adult components. Adult-specific axial skeleton characteristics included apparent fusion of centra and loss of the neural arches of the first two vertebrae, formation of a complete rib-case frequently including hemal arch attachment sites for the last one to two ribs ( Fig. 2 G), a complete set of elaborate dorsal and ventral posterior processes from the centrum ( Fig. 2 G,H) and apparent fusion of the two posterior-most centra (data not shown).
What is the post cranial skeleton?
The post-cranial axial skeleton is derived from somitic mesoderm but the nature of the segmental relationship between these structures remains obscure. The vertebral column develops from sclerotome, a mesenchymal cell population derived from ventral somite. Sclerotome cells that will contribute to the vertebral column move to surround axial midline structures, condense and differentiate as chondrocytes, thus forming a cartilaginous skeletal framework that later is replaced by bone. The somites themselves are segmentally repeating units of paraxial mesoderm. The segmental register of the ‘somite column’ and vertebral column are offset, a fact recognized from the early days of modern embryology (see Remak, 1855) (reviewed by Brand-Saberi and Christ, 2000 ). Experimental studies of somite contribution to the vertebral column has been confined to avian embryos in which sclerotome comprises a major part of the somite. The overall picture that has emerged is that each somite contributes, essentially without significant AP dispersal, to adjacent body structures including sclerotome-derived vertebral components ( Fig. 1 B) ( Beresford, 1983; Aoyama and Asamoto, 1988; Bagnall et al., 1988; Lance-Jones, 1988; Ewan and Everett, 1992; Aoyama and Asamoto, 2000; Huang et al., 2000a ). The original boundary between somites ultimately aligns near the midline of the adjacent vertebral segment, a phenomenon commonly referred to as resegmentation (reviewed by Brand-Saberi and Christ, 2000; Christ et al., 2000; Saga and Takeda, 2001 ).
What is the importance of the vertebral column?
Despite the importance of the vertebral column in defining the overall vertebrate body plan, we still lack a comprehensive understanding of the genetic and cellular interactions that control the size, shape and number of its elements. Investigating sclerotome development from its somitic origins through its differentiation into intricately patterned axial skeleton derivatives provides insight into mechanisms underlying vertebrate segmentation and AP patterning, as well as evolutionary variation among vertebrate species.
What are the ribs and vertebrae?
The vertebral column and ribs that make up the post-cranial axial skeleton have a metameric organization. The functional segmental unit of the vertebral column, the vertebra, is composed, in the simplest terms, of the vertebral body or centrum that develops around the embryonic notochord, dorsally extending neural arches and spines, ventrally extending hemal arches, attachment sites for ribs, and various other decorations of these regions ( Fig. 1 A). These vertebral column elements are distributed in a region-specific manner along the AP axis. Based on such attributes, vertebrae can be grouped into common types, the relative number of each type providing the so-called vertebral or axial formula for a given vertebrate. For example, tetrapods generally have a characteristic number of cervical, thoracic, lumbar, sacral and caudal vertebrae; however, in other vertebrates the vertebral types are less easily categorized (see Romer, 1956 ).
How many clones are in a sclerotome?
Sclerotome progenitors were labeled at 18 h and clone size assayed at 3 d. The fluorescent label often became faint and punctate, precluding a rigorous analysis of sclerotome cell proliferation, although clone size appeared to range between two to six cells. We recorded the distribution of labeled sclerotome cell progeny along the dorsoventral axis in 13 embryos. From a total of 55 cells counted at 3 d, 44 were located dorsal to the notochord or dorsal or lateral to the neural tube, nine were located ventral to the notochord and two were located lateral to the notochord.
What are somites made of?
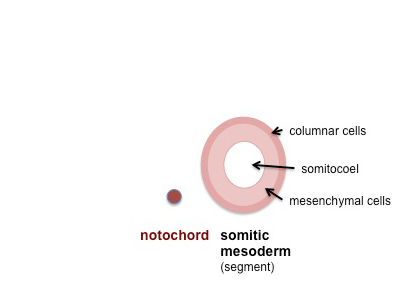
The newly formed somites comprise a sphere of columnar epithelial cells and a central cavity, the somatocoel, containing mesenchymal cells. Early somites are characterized by the expression of the Pax3 gene. Following somite formation, however, expression of the gene is downregulated in the ventral half of the somite epithelium and in the somatocoel cells, whereas it persists in the dorsal half of the somite. The ventral medial cells of the somite subsequently undergo mitosis, lose their epithelial characteristics, and become mesenchymal cells again. This epitheliomesenchymaltransition of the ventral part of the somite is preceded by the expression of Pax1 in the somitic ventral wall and somatocoel cells as it signals the beginning of sclerotome formation. Mutations in Pax1 affect sclerotome differentiation, as reported with different mutations in the undulated ( un) locus ( Balling et al., 1988 ). Successful dorsoventral compartmentalization of somites ultimately leads to the development of the sclerotome ventrally and the dorsally located dermomyotome.
How are somites formed?
During vertebrate development, somites form as paired blocks of mesoderm flanking the notochord and neural tube, giving rise to all skeletal muscles of the body, the axial skeleton, and part of the dermis (McGrew and Pourquiè, 1998 ). Somites are a common feature of the segmented vertebrate body plan and are formed by segmentation of the presomitic mesoderm (PSM) into somites by a clock and wave front mechanism ( Cooke and Zeeman, 1976 ). Somite segmentation is achieved by the formation of a border separating the newly formed somite from the PSM ( Fig. 6.2 F). Differences between the TST of the PSM and the forming somite were originally thought to be involved in border formation and somite separation. However, simulation studies showed that given the high bulk viscosities of the somitic tissues, differences in TST measured in vitro would not suffice to drive these processes ( Damon et al., 2008; Grima and Schnell, 2007 ).
What is the mechanism of somitogenesis?
Somitogenesis has long been known to be driven by mechanisms intrinsic to the presomitic mesoderm. Although many of the elements of the network have been identified, their interaction and their influence on segmentation remain poorly understood. Although mathematical models have been proposed to explain the dynamics of subsets of the network ( Baker et al., 2008; Cinquin, 2007 ), the mechanistic bases remain controversial. In general, two distinct molecular pathways have been implicated in vertebrate segmentation. The first is referred to as the oscillator or “segmentation clock” ( McGrew et al., 1998; Palmeirim et al., 1997; Pourquie, 1999 ). This clock corresponds to a molecular oscillator identified on the basis of rhythmic production of mRNAs for the basic helix-loop-helix (bHLH) Notch target genes mHes1, mHes7, the glycosyltransferase Notch modifier Lunatic fringe ( mLfng ), the Notch ligand Delta-like 1 ( Dll1 ), as well as genes in the Wnt signaling pathways ( Aulehla et al., 2008 ), and also the fibroblast growth factor (FGF) pathway ( Dequeant et al., 2006 ). It is the coordinated regulation of these three pathways that underlies the clock oscillator although it is not known, in any species, which set of genes “drives” the oscillations. Nevertheless, the expression of these genes appears as a wave that arises caudally and progressively sweeps anteriorly across the presomitic mesoderm ( Fig. 2 (A). Although a new wave is initiated once during the formation of each somite, the duration of the progression of each wave equals the time to form two somites. This wave does not result from cell displacement or from signal propagation in the presomitic mesoderm but rather reflects intrinsically coordinated pulses of Lunatic fringe expression.
What are the genes that are involved in somite formation?
A key player in somite border formation is Eph–Ephrin signaling. Eph receptors and Ephrin ligands are expressed in opposing cells at the forming border and are thought to trigger border formation by mediating cell repulsion ( Durbin et al., 1998; Watanabe et al., 2009 ). Other genes that are expressed at the forming somite border and required for somite border formation and segmentation in zebrafish and Xenopus, are N-cadherin ( Warga and Kane, 2007) and PAPC ( Kim et al., 1998 ). Whether any of the genes function in somitogenesis by regulating differential cell adhesion, cortical tension, or other cell properties implicated in cell sorting is not yet fully clear.
What is the segmental nature of the vertebral column?
The segmental nature of the mature vertebral column reflects its origin from the mesenchymal somites of the early embryo. During the fourth week, the ventromedial wall of each somite separates into a mass of diffuse cells, forming the sclerotomes from which the vertebrae originate. 4 The cells of the dorsolateral part of the somite make up the dermomyotome. The dermatome or skin plate lays the foundation for the dermis, and the myotome or muscle plate is the forerunner of striated muscle. This differentiation becomes clinically significant to sclerotomal and dermatomal pain patterns and their origin, corresponding to the development of the spinal nerves directly opposite their respective somites.
What is the process of forming a somite?
Somites begin formation as discrete clusters of mesenchymal cells that undergo segmentation in a cranial to caudal progression beginning around the third week of intrauterine life ; this stage is termed “compaction.” A single somite can be described as having six faces, like a cube, with each facet having a slightly different fate. Further, the position along the embryo can alter the developmental fate of the somite. During this stage, mesenchymal cells multiply rapidly, enclosing the notochord ventromedially as segmented sclerotomes. (See Figure 2-4A .) These will give rise to the bones, joints, and ligaments of the spinal column. Sclerotomal cells later migrate dorsally around the spinal cord, forming the neural arches. The mesenchymatous models in the third stage produce a cartilaginous vertebral column. Single mesenchyme cells will differentiate into chondroblasts when they are maintained in a rounded configuration. In the fourth stage of spinal development, ossification of the cartilaginous vertebral column occurs. 4
What are the signaling molecules in newly formed somites?
Figure 3.27. A simplified scheme of signaling molecules in newly formed epithelial somite. Shh (ochre dots), produced by notochord (Nc) and floor plate, acts on the ventral domain of newly formed epithelial somites, inducing sclerotome, and also on the dorso-medial domain, inducing medial dermomyotome. Wnt1 (red dots), produced by dorsal neural tube (NT), acts (with Shh) on the dorso-medial domain of newly formed somites (Sm), where Myf5 expression is observed soon after and epaxial progenitors are specified. Wnt7a (blue dots), produced by dorsal ectoderm (DE), acts on the dorso-lateral domain, where hypaxial progenitors are specified. BMP4 (brown polygons), produced by lateral mesoderm (LM), prevents MyoD activation and early differentiation in the lateral domain of somites. Its action is counteracted by direct binding of Noggin (green triangles) produced by dorsal neural tube.